标题: 植物相关芽胞杆菌感知假单胞菌铁载体调节次生代谢物
单位:Microbial Processes and Interactions Laboratory, Terra Teaching and Research Center, Gembloux Agro-Bio Tech, University of Liège, Gembloux, Belgium
发表日期:2022年11月10日
摘要
- Bacillus velezensis被认为是植物相关杆菌的模式种,为其宿主提供保护,如防止植物病原体。可能与其广泛分泌的特定和广泛的次生代谢物质有关;
- 代谢物库已经在遗传和化学上得到了很好的定义,但关于它在自然条件下如何表达,特别是它如何在竞争性根际生态位的种间相互作用中被调节,还有待探索;
- 作者在这篇文章中表明:B. velezensis可以感知假单胞菌并动员自身大部分代谢,竞争土壤生态位。
- 这种代谢物变化反映了一种多模式的防御策略,因为它包括具有广泛抗生素活性的聚酮(polyketides)和环状细菌素(bacteriocin amylocyclicin),以及有助于生物膜形成和增强运动性的表面素脂肽(surfactin lipopeptides);
- 假单胞菌次生铁载体pyochelin是一种信息化学物质,通过一种独立于铁胁迫的机制触发B. velezensis的反应。说明:B. velezensis依赖于这种螯合物感知来准确识别竞争对手,是铁载体介导的相互作用的一个新方面。
Plant-associated Bacillus mobilizes its secondary metabolites upon perception of the siderophore pyochelin produced by a Pseudomonas competitor
介绍
由于根系不断渗出,土壤中营养丰富的根际被认为是微生物丰度和多样性最丰富的栖息地之一。细菌和真菌在较小程度上是最主要的形式,但其他微生物,如古菌、卵菌、线虫和原生生物也在根瘤菌群功能和植物健康方面保持重要功能。根际因此代表了种间细菌相互作用的特别的生态位,无论它们是合作的还是竞争的。
由于先进的基因组学、分析和成像技术,在破译这种竞争性相互作用的分子基础方面取得了实质性进展。它涉及信号干扰、接触依赖性传递系统释放毒素,还涉及挥发物和小尺寸可溶性代谢物的产生,如群体感应分子、亚抑制浓度的抗生素和降低铁可利用性的铁载体。为了响应竞争对手的感知,细菌调整其表型并刺激次生代谢物的产生,这些代谢物作为抗菌剂或影响发育性状,如生物膜形成、运动性和产孢。这些增加的代谢物在大多数情况下对应于已知的化合物或结构衍生物,但物种间的相互作用也可能解开隐藏分子的合成。
几种属于枯草芽孢杆菌群的芽孢杆菌是根瘤菌群中普遍存在,包括B. velezensis(植物相关模式菌株),已知在促进生长和防止植物病原体方面为其宿主提供益处。B. velezensis与枯草芽孢杆菌群的其他物种的区别在于将其基因组的10%以上用于负责合成特殊代谢物的基因簇。这种次级代谢组具有化学多样性,包括挥发性化合物、萜烯、非核糖体二肽、环脂肽(CLPs)和多酮(PKs),也包括核糖体合成和翻译后修饰的抗生素和细菌素。目前,对芽孢杆菌生物活性次生代谢物的研究主要集中在植物病害生物防治方面的生物活性表征,描述了它们直接抑制植物病原体的作用,以及它们作为宿主免疫激发子的活性,从而增强了对感染的全身性抵抗。然而,从生态学的观点来看,这些化合物中的一些保留了其他功能,有助于生产细菌在自然环境中持续存在。然而,对于B. velezensis,与其他物种一样,作者对次生代谢物的生态相关性的了解仍然有限,并且在多营养相互作用中的许多发育和信号功能显然仍有待发现。
对于其他细菌物种,人们认为微生物相互作用是调节芽孢杆菌产生生物活性次生代谢物的主要生物因子。这已经在几项研究中得到了证实,这些研究报道了B. velezensis在感知一系列土壤真菌和卵菌时增强了抗真菌CLPs iturin和fengycin的产生。芽孢杆菌也与植物有益的丛枝菌根真菌相关,但对分子信号知之甚少,也没有迹象表明其可能调节次生代谢物。最近的一些报道描述了土壤杆菌在面对细菌竞争对手时如何调节自己的行为,但几乎只描述了对发育性状的影响,如社会运动、生物膜形成和产孢或适应策略。到目前为止,只有少数论文报道了芽孢杆菌在细菌相互作用下可能调节次生代谢物的产生。在与细菌植物病原体共同培养时,已经观察到一些负责合成表面素,杆菌素或iturin的基因的表达增强。然而,在这些研究中,以及在芽孢杆菌与其他植物有益物质相互作用的背景下,没有报道这种转录刺激与相应代谢物高产之间的相关性。
在这项工作中,作者主要想评估B. velezensis在感知细菌竞争对手(类似根际生态位分离)时调节其次级代谢组的潜力。作者研究了一个简化的二元系统中的成对相互作用,足以揭示细胞-细胞信号调控。作者在非接触的环境中进行了大多数实验,以研究远距离的交叉对话,即由可扩散化合物的感知介导。本文使用的无柄假单胞菌(Pseudomonas sessilinigenes)属于荧光假单胞菌群,被选为根瘤菌群中常见的植物有益但竞争激烈的挑战者。它还保留了合成多种生物活性次级代谢物的潜力,从而可以研究两种物种之间的化学相互作用。通过结合分子和分析方法以及突变方法,作者观察到B. velezensis通过化学感知假单胞菌的竞争对手来调动其次级代谢组的很大一部分。由于它包括几种聚酮和细菌素链环素,这种代谢物反应与增强的全球抗菌潜力相关。相互作用也会刺激表面素的产生,这可能有助于根际适应性,因为它有利于生物膜的形成和运动[33]。作者还想破译导致诱导的机制,并确定假单胞菌次级铁载体(对外)pyochelin (E-PCH)是芽孢杆菌特异性感知的信号。这为铁载体在种间化学对话信号中的作用提供了新的线索。在微生物相互作用中,这种螯合剂感应现象对芽孢杆菌的潜在益处也进行了讨论。
结果
B. velezensis在感知假单胞菌代谢物时调节其次级代谢组
使用遗传上可适应的天然根际分离物GA1作为B. velezensis的代表。事实上,基于基因组挖掘,它保留了产生全套生物活性次生代谢物的潜力,这些代谢物通常由该物种的成员形成。在所谓的模拟分泌物培养基中生长时,整个代谢物范围的高效生产证实了这一点,反映了茄科植物根系释放的主要碳源中的含量(图S1)。它包括三个clp家族(surfactin, iturin, fengycin),二肽杆菌素,铁离子杆菌素和三个多酮家族(bacillaene, difficidin,和macrolactin)。此外核糖体合成和翻译后修饰肽,如amylocyclicin和amylolyysin也存在于GA1基因组中。然而,基于UPLC-qTOF质谱测定的精确质量,这些化合物无法在培养液中可靠地检测到。选择了与植物相关的分离物P. sessilinigenes CMR12a作为主要的相互作用伙伴,因为它能产生多种次级代谢产物,如铁载体pyoverdin和enantio-pyochelin(E-PCH),抗生素吩嗪(phenazine)以及两种结构不同的CLPs, sessilins和orfamides。这些化合物是在酪氨酸培养基中培养的,通常用于假单胞菌在铁限制条件下的培养(图S2)。
作者评估了GA1在与CMR12a相互作用时调节次生代谢物产生的潜力,这种作用避免了由于半固体基质中的扩散限制或由于不可渗透生物膜结构的形成而可能产生的干扰。因此,通过对添加/不添加含有CMR12a在酪胺酸培养基中产生的代谢物的无菌发酵液(下面称为粗提物,2% (v/v))的GA1液体培养基生长来进行检测。作者首先采用非靶向UPLC-MS方法,比较了指数期开始和结束及稳定期的三个不同时间点下GA1在两种条件下分泌的代谢物。使用MZmine 2进行数据处理使作者能够检测出在交互作用下产生的特征和相应的天然产物差异。结果显示,添加假单胞菌粗提取物后,三种次级代谢物的生长阶段依赖性差异积累(图1a,图S3为GA1生长动力学)。表面素(surfactins)的生成在指数期早期显著增加,杆菌烯(bacillaene)或二氢杆菌烯(2H-bae)在指数期向稳定期过渡时受到刺激,铁载体杆菌素(siderophore bacillibactin)的合成在稳定期增强(图1a)。因此,增强效应的幅度和时间根据分子而变化,很可能是由于特定的转录调节。添加CMR12a粗提取物不影响其他化合物如fengycins、iturins、大乳酸素macrolactins和杆菌素bacilysin的产生(图S4)。接下来,作者开始使用优化的UPLC-MS方法检测生长20小时后的整个天然产物范围,以靶向非核糖体化合物。它证实了(二氢)芽胞杆菌烯、表面素和芽孢杆菌素的不同积累,但也显示添加CMR12a提取物后,艰难梭菌素difficidin 或其氧化形式(仅在ESI阴性模式下检测到)的产生显著增强(图1b,化学结构见图1c)。作者在全球范围内观察到假单胞菌CMR12a提取物对其他B. velezensis生物防治菌株(如S499、FZB42和QST713)次生代谢物产生的类似影响(图S5),表明这种相互作用介导的代谢物反应在该物种成员中是保守的。
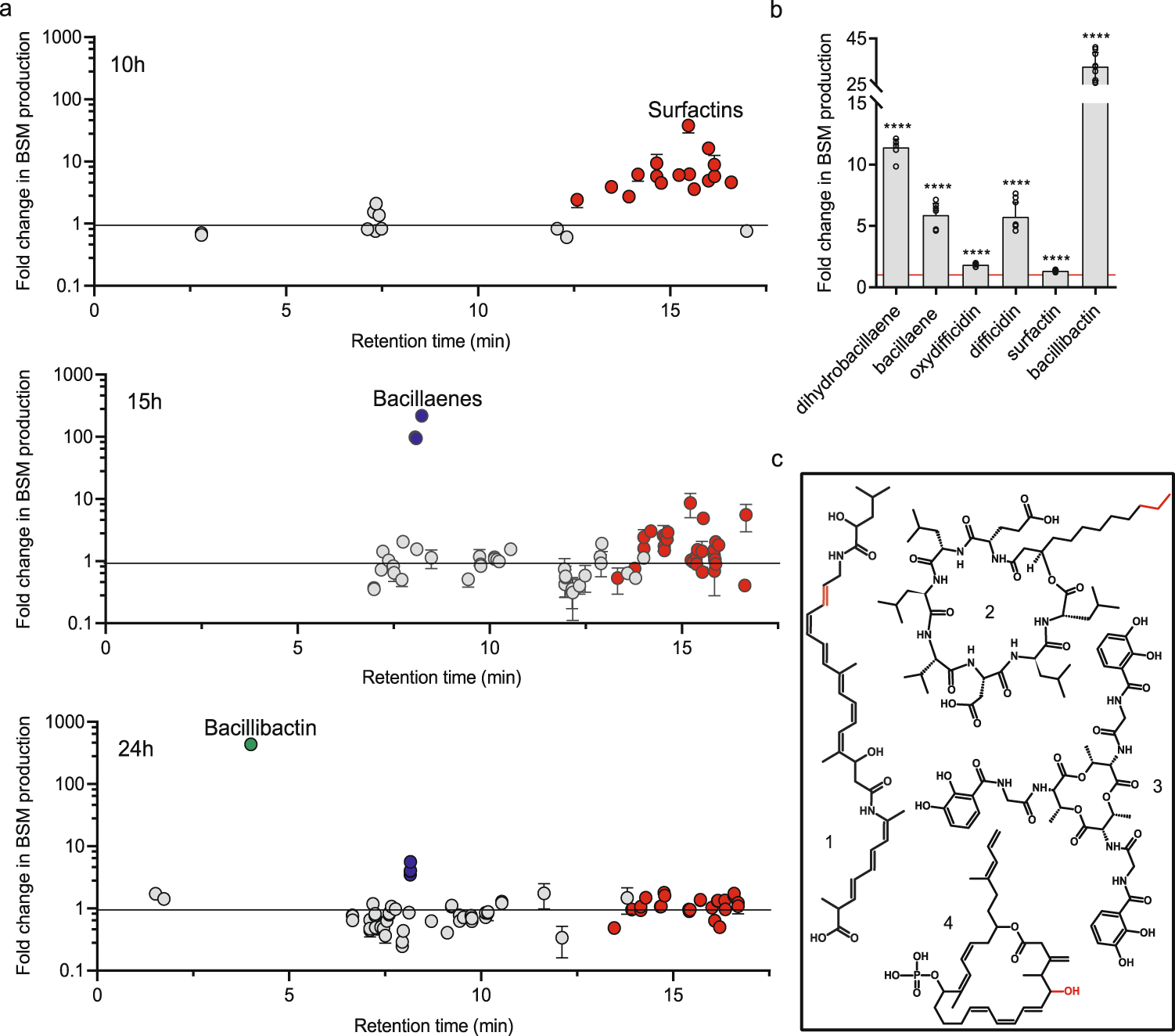
a 添加CMR12a无细胞上清液对指数期早期(10 h)、指数期晚期(15 h)和稳定期(24 h) GA1代谢组的影响。每个点代表检测到的一个特征。红色、蓝色和绿色的点分别代表了表面素、杆菌素和杆菌素的特征(变体或加合物),而灰色的点代表了GA1代谢产物的其余部分。与对照培养物(未添加GA1培养物)相比,生产过剩数据表示为每OD600的峰值面积折叠变化。误差条表示标准差(n = 3)。
b 与未添加CMR12a无细胞上清(2% (v/v))的培养物相比,添加CMR12a无细胞上清后GA1生物活性次级代谢物产量增加了2倍(倍数变化= 1,红线)。数据的计算方法如a所示。均值由三个独立实验(n = 9)三次重复得到的数据计算。使用Mann-Whitney检验计算统计显著性,其中“”表示p < 0.0001的显著差异。c过量生成的bsm结构,红色部分为可变部分,解释了大多数化合物(1:(二氢)杆菌烯,2:表面素;3: bacillibactin;4:(氧)difficidin)。
生物活性次级代谢物刺激导致抗菌潜力增强
PKs(如:(二氢)芽胞杆菌烯、(oxy)艰难梭菌素和大乳酸素)是芽孢杆菌的次级代谢物之一,因其对多种细菌的抑制活性而得到最好的描述。作者推测增强的抗菌活性可能是GA1代谢物对CMR12a产物反应的直接结果。这是通过测试相互作用产生的芽孢杆菌提取物观察到的,用于分别作为革兰氏阴性和革兰氏阳性植物致病菌的代表,对Xanthomonas campestris和Clavibacter michiganensis的生长抑制。在这些条件下,假单胞菌无细胞上清(0.08% (v/v)(0.04×0.02))对病原菌生长没有影响,这显然是由于培养基中代谢物的浓度很低。为了确定每种代谢物在细菌抑制中的具体作用,我们生成并测试了一系列GA1敲除突变体,包括Δsfp突变体,该衍生物特异性抑制4’-磷酸泛酰巯基乙胺基转移酶,该酶对于PK和非核糖体肽生物合成机制的正常运作至关重要(即负责PK, CLPs和bacillibactin的合成)。Δsfp提取物中抗黄单胞菌活性的完全丧失表明了这些化合物的关键作用(图2b),并排除了其他已知具有抗菌活性的分子的可能参与,如杆菌素或核糖体合成和翻译后修饰的肽(图S6a)。在单个化合物合成中被特异性抑制的突变体的功能丧失,指出了(oxy)艰难梭菌素和(二氢)杆菌烯在抑制X. campestris中的关键作用(图2b)。单独或协同作用,这两种PKs也负责GA1对其他重要的细菌植物病原体的抑制活性,如胡萝卜假单胞菌(P. carotovorum)、A. tummefaciens和R. fasciens,但不参与抑制P. fuscovaginae物种,其中杆菌素可能是活性代谢物(图S6b)。然而,GA1对这里测试的非致病性土壤P. essilinigenes CMR12a和P. protegens Pf-5没有表现出明显的毒性(图S7)。
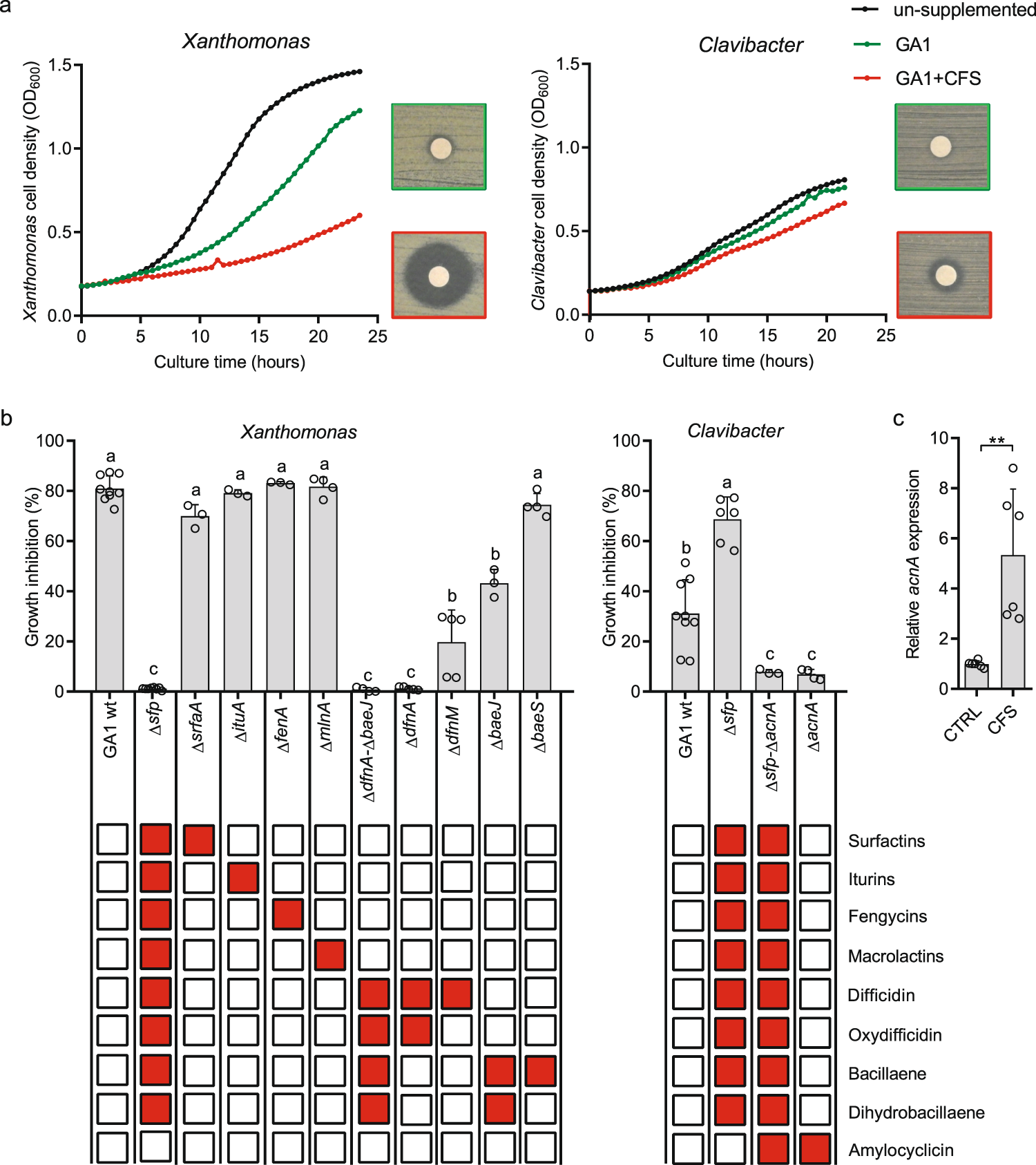
与X. campestris相比,增强的对C. michiganensis的抗生素活性不是由非核糖体产物介导的,如Δsfp突变体的高活性所示(图2b)。与GA1野生型相比,Δsfp突变体对密歇根赤霉病的抗性明显增强。在这个突变体中,原本有效产生的CLPs的产生被取消了。在此基础上,我们预计CLPs合成所需的氨基酸将被保留,我们假设它们可能被重定向到核糖体肽的生产中,包括那些对C. michiganensis有活性的氨基酸,从而导致活性增强。因此,我们从基因组数据和文献推测核糖体合成和翻译后修饰的肽,如amylocyclicin,可能参与了抑制作用。这一假设得到了相应生物合成基因的ΔacnA突变体敲除所观察到的病原体抑制潜力的急剧丧失的支持(图2b)。此外,另一种抗菌肽amyolysin的突变体和不依赖于单肽的非核糖体肽bacilysin没有失去活性(图S6a),这表明amylocyclicin是拮抗Clavibacter的主要代谢物。我们无法提供证据证明培养基中成熟肽的积累更高,但RT-qPCR数据显示,在CMR12a的作用下,GA1细胞中acnA基因的表达被强烈诱导(图2c)。基于基因组挖掘,S499是另一种B. velezensis菌株,它也保留了产生amylocyclicin的潜力,并且在CMR12a产物存在的情况下,该菌株也观察到acnA基因的表达增强(图S8)。
次生代谢物的合成在竞争性根定植中得到增强
因此,这些体外数据说明了在酪氨酸培养基中生长的浮游细胞分泌的一些假单胞菌化合物如何调节B. velezensis的次级代谢组。然而,除了pyoverdine外,所有CMR12a代谢物都很容易在模拟渗出液的培养基中产生,并且当细菌定植在番茄根部时也会形成大量的CMR12a代谢物(图2)。S1和S2)。因此,我们接下来用22°C生长的番茄植株进行了植物实验[48],以测试CMR12a存在时根共定植时GA1的代谢物刺激。当单独接种时,两种分离株都能有效地定植根,但在竞争性根入侵时,CMR12a超过GA1,与单接种植株相比,其形成的种群明显减少(图3a)。这可能是由于假单胞菌更高的内在生长速率和/或更好地利用渗出物,但也可能是由于产生了抗菌剂,如最近被描述为具有抑制贝氏杆菌活性的脂肽sessilin[36]。由于这些减少的群体,我们无法可靠地检测到共接种植物根际提取物中的所有GA1代谢物。然而,在与CMR12a共接种的GA1细胞中,与单独接种相比,观察到负责合成surfactin、(dihydro)bacillaene、(oxy)difficidin和amyocyclin的基因簇的表达显著增强(图3b)。这表明浮游GA1细胞观察到的代谢物刺激也发生在竞争性定植的背景下,细菌形成生物膜,完全以根分泌物为食,显然与挑战者的菌落密切接触。
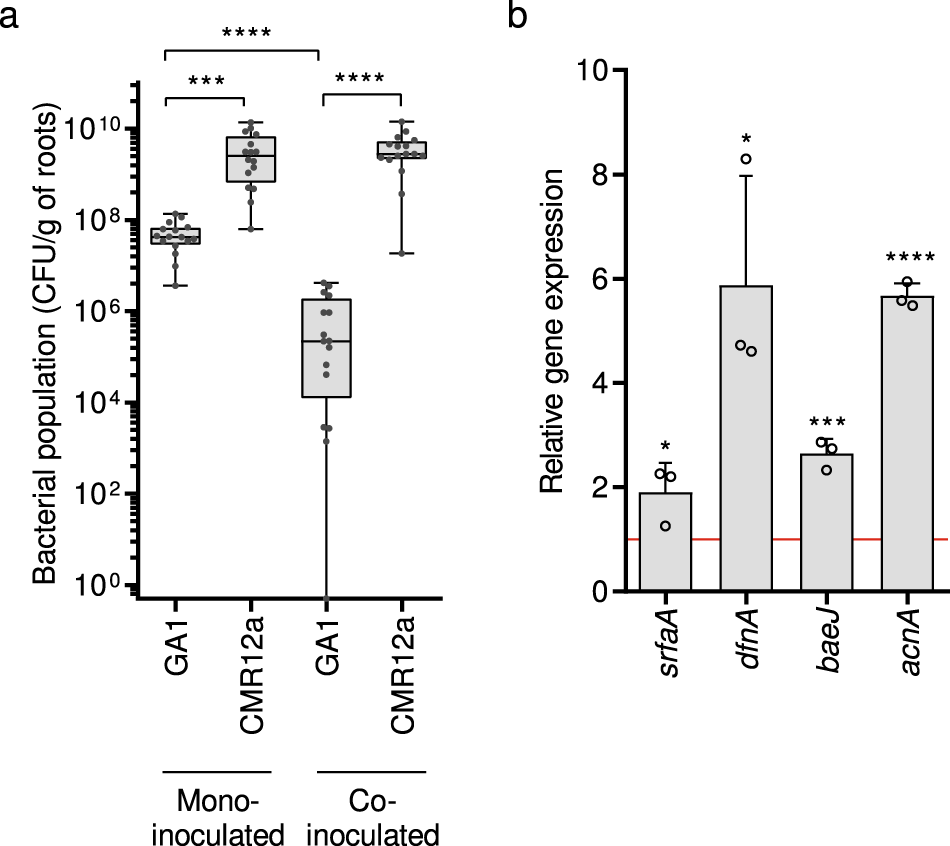
图3:竞争定植实验支持了植物中芽孢杆菌-假单胞菌相互作用中生物活性次级代谢物的作用。
a 分别接种或共接种番茄幼苗,接种后3天从根部恢复GA1和CMR12a细胞群。箱形图是根据4个独立试验的数据创建的,每次试验至少涉及4株植物(n = 16)。采用Mann-Whitney检验分析处理间的统计学差异,“****”和“***”分别代表p < 0.0001和p < 0.001的显著差异。
b 在植物(番茄根部3 dpi) GA1中,分别负责合成表面素、(氧)艰难梭菌素、(二氢)芽胞杆菌素和支链霉素的sfraA、dfnA、baeJ和acnA基因的相对表达。图表显示了三个生物重复(n = 3)计算的平均值和±SD,每个重复涉及六种植物。红线中的Fold change = 1对应单独接种GA1在根上的基因表达,作为对照条件。共定殖组与对照组数据采用t检验进行统计学比较(* p < 0.05; *** p < 0.001;**** p < 0.0001)。
脓毒杆菌素(pyochelin)是芽孢杆菌感知的假单胞菌信号
接下来,我们希望进一步研究这种化学对话的分子基础,并确定由CMR12a分泌的化合物,这些化合物被芽孢杆菌细胞感知以触发抗菌剂的产生。为此,我们使用2H-bae作为芽孢杆菌反应的主要指标,因为考虑到迄今为止尚未报道对bacillibactin的直接毒性活性,2H-bae代表了具有抗菌活性的最高增强代谢物。
我们首先在C18固相萃取筒上对CMR12a无细胞上清进行生物活性引导分离。结果显示,只有含有pyoverdine和/或E-PCH的提取物显示出一致的PK触发活性(图4a)。从CMR12a培养物中制备的无细胞上清液在添加了Fe3+的酪胺酸培养基中活性急剧降低,从而抑制了它们的产生,从而支持了铁载体的可能参与(图4b和S9)。接下来,我们比较了从CMR12a敲除突变体的培养物中获得的无细胞上清液的触发潜力,该突变体特别缺乏不同的已鉴定代谢物,但在其他代谢物的产生中没有被破坏。正如预期的那样,只有在E-PCH产生中受损的突变体的提取物在PK诱导潜力方面受到显著影响(图4c)。为了证实两种铁载体的这种差异效应,我们随后以与假单胞菌无细胞上清液中测得的浓度相似的浓度独立测试了HPLC纯化的化合物。数据显示,与CMR12a产生的主要绿脓杆菌定异构体相比,E-PCH的PK触发活性高得多(图4b)。
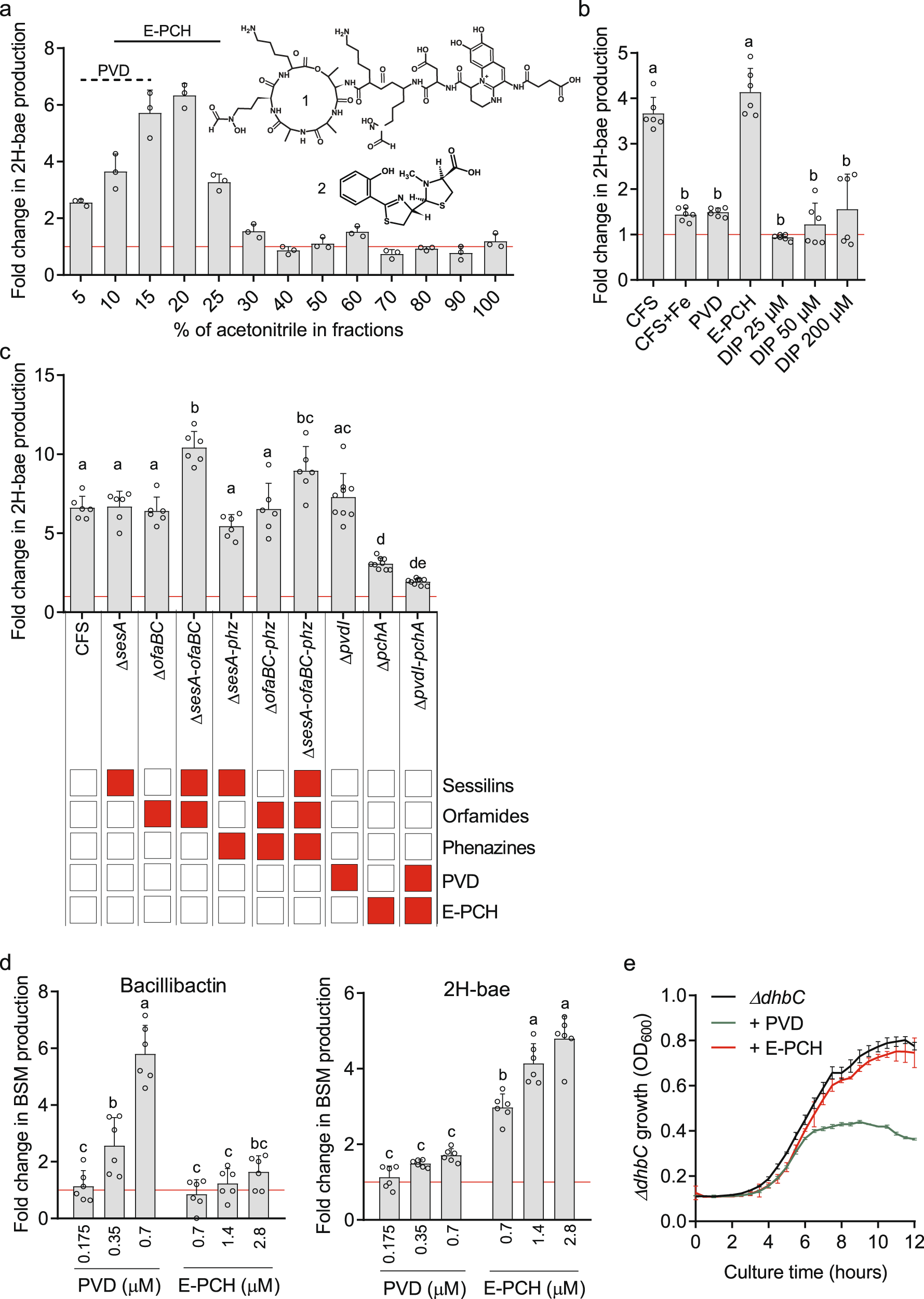
a CMR12a无细胞上清(CFS)在C18墨盒上进行固相萃取处理,根据化合物的疏水性,随着乙腈-水比(以ACN % (v/v)表示)的增加,逐步洗脱化合物。所有馏分均采用UPLC-MS分析,含有大量pyoverdine (PVD) (1: PVD结构)和/或对映体pyochelin (E-PCH结构)(2:E-PCH结构)的馏分分别用虚线和实线标记。通过UPLC-MS(峰面积)对处理过的培养物中每OD600的化合物进行相对定量,与未添加的对照比较。
b 添加0.35µM纯PVD、1.4µM纯E-PCH、2% (v/v) CMR12a无细胞上清(CFS)、补铁培养(CFS + Fe) CMR12a CFS和不同浓度的铁螯合剂2,2′-dipyridyl (DIP)后,二氢杆菌烯(2H-bae)产量的差异。在GA1培养物中添加CMR12a野生型和各种突变体CFS (2% (v/v))对2H-bae产量的影响。在CMR12a突变体中特异性抑制的代谢物用红框表示。a. d .纯PVD和E-PCH对bacillibactin和2H-bae产量的剂量依赖性影响。GA1培养物中添加指定浓度的hplc纯化的CMR12a铁载体。折差计算如a所示。b、c和d表示的数据是两个独立实验(n = 6)中三个重复培养计算的平均值和±SD,其中不同字母表示2H-bae和bacillibactin的差异有统计学意义(ANOVA和Tukey’s HSD检验,α = 0.05),对照组(即未添加GA1培养)属于c组。
e 添加纯PVD和e - pch对杆菌杆菌素合成抑制的GA1 ΔdhbC突变体生长的影响。添加CMR12a铁载体的最终浓度与添加2% (v/v) CMR12a CFS(0.35µM PVD和1.4µM E-PCH)的最终浓度相似。平均值和±SD来自三个重复。详细统计分析见图S10。
在另外的剂量依赖性试验中,我们观察到补充pyoverdine导致培养基中的铁限制,这是由GA1感知的。这可以从GA1野生型中铁载体bacillibactin的产量显著增加(图4d)和添加pyoverdine后抑制bacillibactin合成的ΔdhbC突变体的生长减少(图4e和S10)来说明(图4e和S10)。我们的基因组检测显示,B. velezensis GA1具有多种转运蛋白,可以吸收外源的铁载体(表S3),与枯草芽孢杆菌中发现的转运蛋白相似。然而,数据表明pyoverdine(以铁的形式)不能被GA1吸收。pyoverdine作为强螯合剂,清除外界介质中大部分有效铁,引起GA1细胞铁应激。这导致在转录抑制因子Fur介导的过程中GA1铁载体bacillibactin的产生增强。pyoverdine或2,2 ‘ -dipyridyl (DIP)化学螯合剂的加入,都没有被芽胞杆菌内化和使用,也会导致2H-bae合成的轻微但不显著的刺激(图4b, d)。我们假设,正如之前报道的那样,由于外源性铁载体/螯合剂的存在,2H-bae的低增加是对铁限制的整体反应的一部分。在GA1中,2H-bae的合成受全局调控因子Fur的控制,该调控因子参与铁稳态并控制杆菌杆菌素的产生,因为在Fur缺失突变体中,2H-bae的重新合成显著增强(图S11a)。这种调控很可能是间接的,因为我们无法在bae操纵子的启动子区域找到任何Fur结合位点(Fur-box)(表S4)。
与pyoverdine相比,对铁亲和力低得多的次级铁载体E-PCH的加入强烈刺激2H-bae的产生,但不激活bacillibactin的合成(图4d),并且在使用的浓度下不影响ΔdhbC的生长(图4e和S10)。因此,E-PCH不会通过引起铁限制作用于GA1细胞,这一事实进一步支持了E-PCH在ΔfurR突变体中对2H-bae合成的刺激是保守的,对生长没有影响(图S11a, b)。也就是说,据报道,E-PCH通过引起氧化应激和损伤来保留一些抗生素活性,如在大肠杆菌中报道的。然而,E-PCH对GA1野生型和ΔdhbC突变体没有表现出任何毒性,也不会显著影响参与芽孢杆菌氧化应激反应的主要调节因子中被抑制的ΔperR突变体的生长(图S11b)。此外,在ΔperR突变体中也保持了2H-bae的持续增加(图S11a),进一步表明E-PCH不会通过作用于细胞内诱导氧化应激,从而支持其被芽孢杆菌细胞视为外源信息化学物质。这种信号传导活性不受分子立体化学的影响,因为我们通过测试从铜绿假单胞菌PAO1中纯化的另一种天然异构体pyochelin (PCH),观察到聚酮生成的类似增加(6.05±1.8倍,n = 4)。
P. protegens菌株如Pf-5和CHA0也被描述为E-PCH生产者。我们还观察到,添加由Pf-5制备的无细胞上清液后,PK产量的增加与CMR12a相似(图S12)。pchE基因编码形成线性三环PCH支架的2-羟基苯基噻唑啉基硫酯中间体的关键酶,这在分类上密切相关的P. protegens和P. sessilinigenes中非常相似(92%的同源性)(图S13)。在文献中,也有关于伯克霍尔德氏菌和链霉菌产生PCH或结构相关化合物的报道。但PchE序列的保守性要低得多(50%同一性,图S13),这表明这些蛋白质具有相似的功能,但可能是独立进化的。
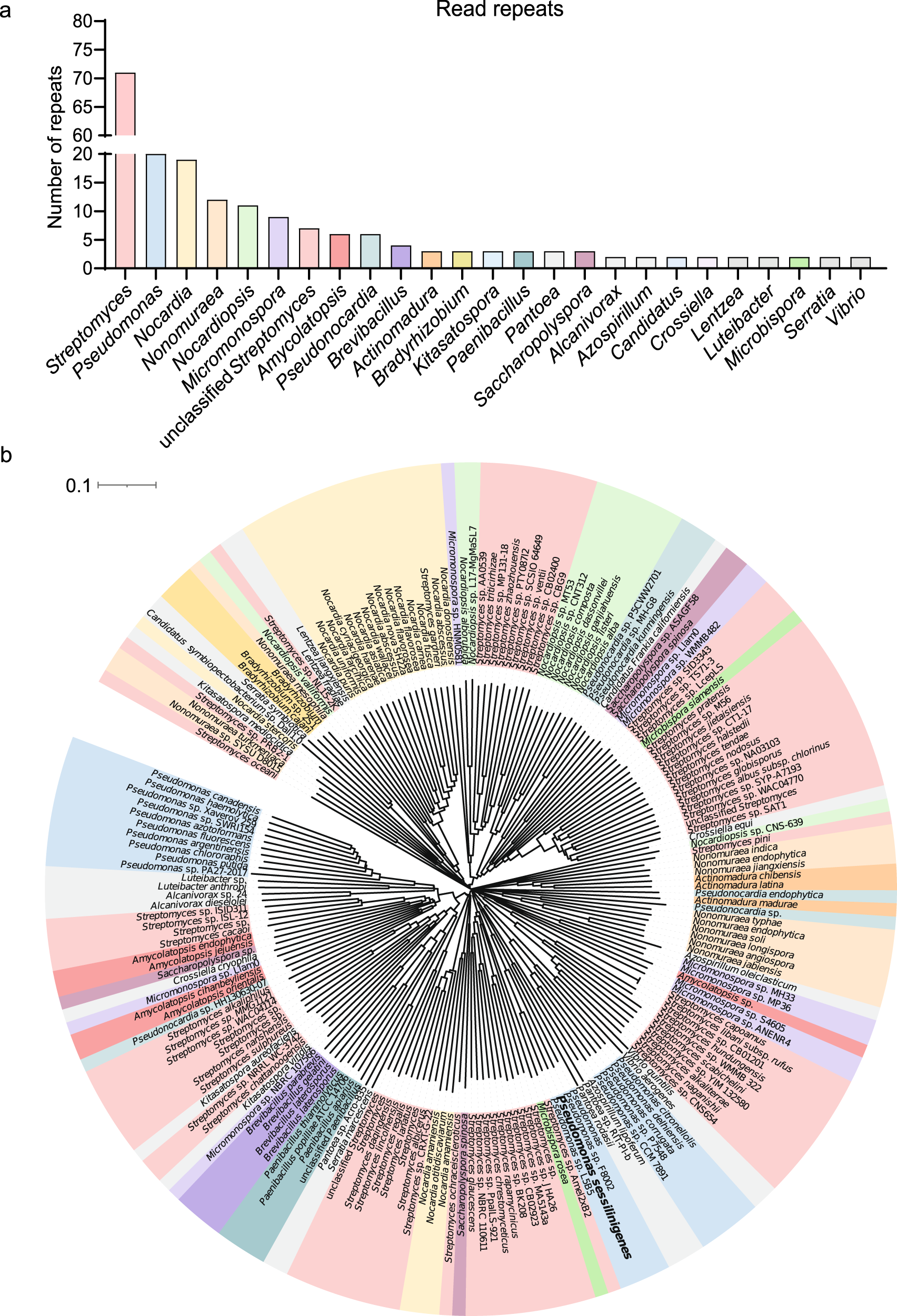
我们还希望在细菌门中搜索由PchE基因编码的PchE蛋白的假定同源物,以评估产生PCH的潜力有多广泛。为了支持该基因和相关化合物的重要性,它首次揭示了在系统发育上遥远的细菌分支中几乎整个序列(>90%)的高度保守性(>40%序列同源性)。对BLAST结果进行过滤后,发现PchE蛋白的假定同源物存在于多种细菌中,包括许多假单胞菌和放线菌,如链霉菌、诺卡菌和小单孢菌(图5a)。为了支持这些数据,有报道称这些放线菌种类以及短芽孢杆菌和弧菌属的成员产生PCH或PCH同源物。所有已鉴定的物种都被描述为土壤栖息细菌,因此可以与B. velezensis在同一生态位中找到。接下来,基于blast结果对253个有代表性的序列进行ClustalW多序列比对,构建了一个Neighbor-Joining系统发育树(图5b)。我们在该树的几个分支中观察到同一属的聚类,这可能表明编码PchE相关蛋白的基因存在密切的类似物,主要是在链霉菌物种中。这表明在非核糖体肽合成酶机制和PCH相关合成化合物的结构上存在一些多样性。