A plant genetic network for preventing dysbiosis in the phyllosphere

Highlight
- 植物叶际phyllosphere,是微生物群落定殖最丰富的环境之一,在全球二氧化碳和氧平衡中起关键作用;
- 植物是否以及如何控制叶际微生物群以确保植物健康?
- mfec四突变(min7fls2efrcerk1)(PTI和MIN7介导的囊泡运输存在缺陷),cad1突变(组成激活细胞死亡缺陷),改变了内生叶层微生物(microbiota),并表现出与生态失调相关的叶片组织损伤(leaf-tissue damage associated with dysbiosis)。
- mfec和cad1突变体的Shannon diversity index和Firmicutes显著低于WT,Proteobacteria被富集;
- 细菌群落移植实验(Bacterial community transplantation experiments)证明,正确组装的叶际细菌群落在叶环健康中具有因果关系;
- PTI、MIN7和CAD1在主要陆地植物谱系中都存在,这可能是陆生植物通过调节该遗传网络控制多样的内生叶际微生物群,保持自身健康的方式。
2016年,密歇根州立大学何胜洋教授团队在Nature上发表的科研成果揭示了一种细菌感染植物的新机制,即细菌除了能抑制植物的免疫系统之外,也会在植物体内产生一种潮湿的环境,加上高湿度的条件,容易引发植物病害。
Result
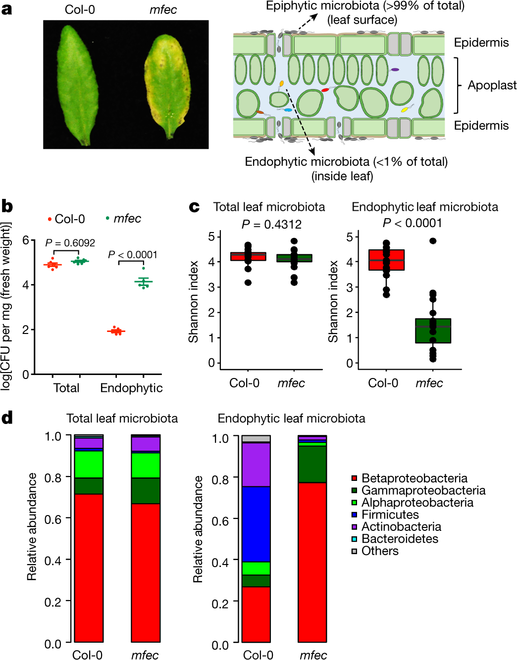 作者团队利用拟南芥四突变体mfec (min7 fls2 efr cerk1)在高湿度的条件下显现出黄化和坏死的病症的特点,通过16S rRNA基因扩增子测序检测了健康叶片(野生型,Col-0)和不健康叶片(突变体,mfec)叶片总微生物组和叶内微生物组,结果表明,健康叶片和不健康叶片的总微生物组的多样性没有显著差异,但是==叶内微生物组的丰度和多样性均有显著的变化==,不健康的叶片内部微生物的多样性降低,从Firmicute-rich的群体转化为Proteobacteria-rich的群体(图 1)。
作者团队利用拟南芥四突变体mfec (min7 fls2 efr cerk1)在高湿度的条件下显现出黄化和坏死的病症的特点,通过16S rRNA基因扩增子测序检测了健康叶片(野生型,Col-0)和不健康叶片(突变体,mfec)叶片总微生物组和叶内微生物组,结果表明,健康叶片和不健康叶片的总微生物组的多样性没有显著差异,但是==叶内微生物组的丰度和多样性均有显著的变化==,不健康的叶片内部微生物的多样性降低,从Firmicute-rich的群体转化为Proteobacteria-rich的群体(图 1)。
接种(inoculated)健康Col-0拟南芥叶际内生的由48个成员组成的群落(SynCom^Col-0^),16S rRNA基因测序分析。
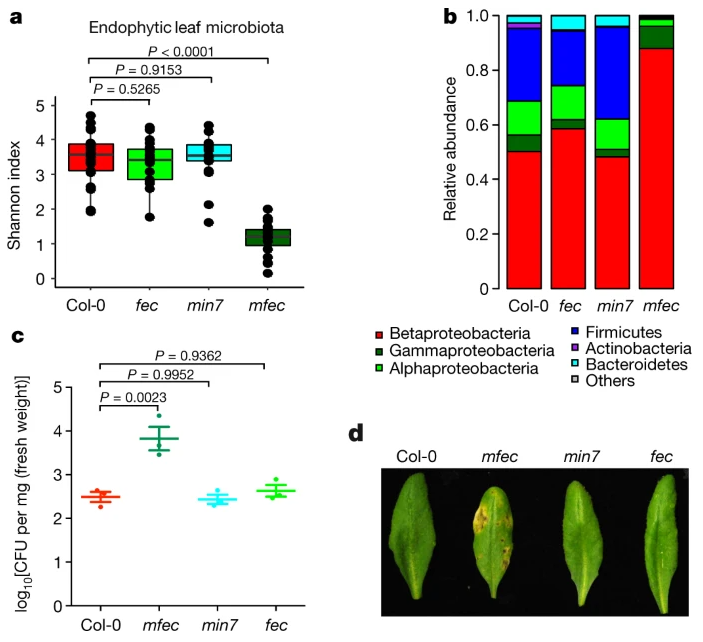
与Col-0叶子相比,mfec叶片中总体多样性显着降低,并且内生细菌群落组成发生了显着变化,但fec(PTI缺陷)或min7(仅在MIN7途径中缺陷)没有发生变化。相应地,只有mfec植物显示出坏死和萎黄病,并且具有较高水平的内生细菌菌群。这些结果表明PTI和MIN7囊泡运输途径在控制拟南芥内生叶微生物区系中的非冗余和必不可少的作用。
mfec植物的组织损伤可能是由植物的一种生态失调引起的→无菌实验:mfec植物在没有微生物群的情况下看起来很健康
由52个成员组成的mfec叶子衍生的内生细菌群落(SynCom^mfec^),improperly assembled
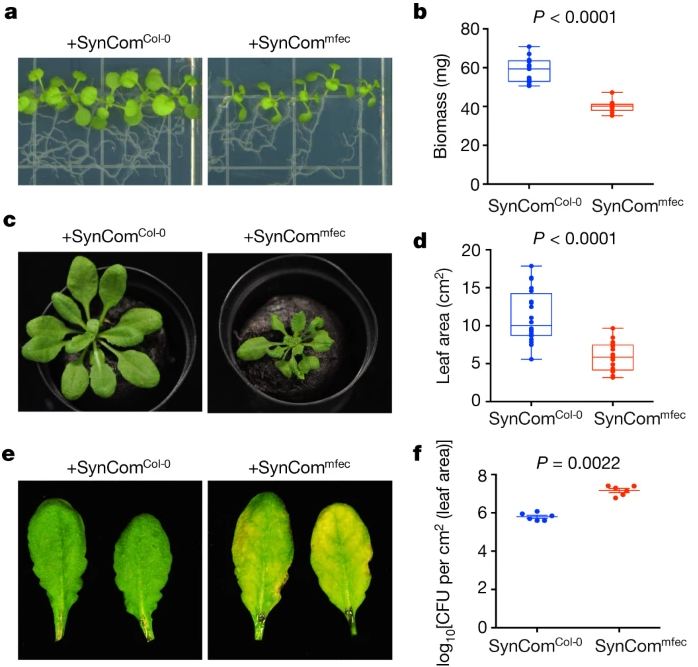
进行了三种类型的功能测定,以严格测试与mfec相关的微生物群是否会导致损害健康的营养不良。
-
在Linsmaier–Skoog琼脂平板测定中,接种SynCom^mfec^的Col-0植物相对于接种SynCom^Col-0^的植物生物量明显减少;
-
当在基于泥炭的GnotoPots中生长时,在SynCom^Col-0^存在下Col-0植物看起来健康,但在SynCom^mfec^存在下表现出不同程度的幼苗发育迟缓和莲座丛大小总体减少;
-
当以每毫升1×10^8^ CFU浓度渗入完全生长并定植的Col-0植物的健康叶片中时,SynCom^mfec^而非SynCom^Col-0^会诱导明显的坏死和萎黄。
此外,在Col-0叶片中,SynCom^mfec^的群落数量比SynCom^Col-0^多。这些综合结果表明,不良生物微生物区系(SynCom^mfec^)足以在野生型植物中产生负面的健康影响,并提供了组装正常的叶片内生微生物区以确保叶环健康的重要性的证据。
研究了mfec植物失去维持内生叶细菌多样性的能力的潜在机制
2116(46×46)个二元相互作用实验显示:大多数厚壁菌门被变形菌门抑制;在正常植物的叶片表面和叶内,各微生物的含量很低,没有达到抑制的浓度和距离;而在mefc突变植物中,显著增加了植物叶片内微生物的生长,于是表现出变形菌门对厚壁菌门的抑制作用。
A framework for microbiota homeostasis
模式触发的免疫和MIN7囊泡运输途径在控制内生叶球微生物中起重要作用,但是不清楚这两个途径是独立作用还是协同作用的。
*ben3*(胞内囊泡运输缺陷,*BIG2*突变)突变体于*mfec*表型相似:都有自然生态失调表型;叶内菌群含量比野生型高;Shannon index低,变形菌门丰富、厚壁菌门减少。ben3的表型是由于cad1^S205F^引起的。
Question
- 在文中,作者在探究mfec突变体表型是否是由于植物营养不良引起时,通过实验证明:在无菌条件下,mfec突变体与野生型无明显差异,而在接种SynCom^Col-0^后表现出营养不良的症状。因此,说明了SynCom^Col-0^解释了mfec宿主的表型。
那这种表型到底是由于广泛的 ==细菌增殖== 引起的还是由 ==厚壁菌门的缺失== 引起的?比较好奇在mfec突变体在无菌条件下接种Syncom^Col-0^中的厚壁菌门后的表型。
相关实验:
三型效应因子相关基因突变的Pto DC3000(非致病、不能使Col-0表现出患病表型)可以使mfec发生变黄等表型(湿度95%),相对负载有50-1000倍差异
→是不是可能其他菌也能有相似的表型?还是需要更多的实验验证。
作者一直强调 ==高微生物多样性== 与 ==植物健康== 的关系,认为高生物多样性与植物健康密切相关,高生物多样性促进了植物健康,植物健康维持了微生物高多样性,我同意这个观点。而我,认为更应该认识到 ==微生物负载== 与 ==植物健康== 的关系。当植物宿主的实际微生物负载超过其所能承受的负载时(增加了营养利用),植物会表现出患病的表型,宿主为了防止这种不平衡的出现,从而通过全身协同作用调控,抑制微生物的生长,维持相对稳定。
- 所有突变体的Total微生物是没有显著差异的
TIps
灭菌营养土2次(121℃,45min,隔一天)
表现病症的叶片中微生物数量比正常叶片高50-1000倍(==微生物增加==和==植物患病==谁是因果?我认为是微生物的增加是因,植物患病是果??)